Dacă o celulă funcționează în condiții anaerobe, produce energie prin transformarea glucozei în lactat și, prin ciclul Cori, elimină acesta din urmă; dacă există oxigen disponibil (deci în condiții de repaus), mai mult de 90% din glucoză este consumată aerob și doar restul de 10%, anaerob. Atunci când este nevoie de mai mult ATP decât ceea ce calea aerobă este capabilă să ofere (de exemplu, atunci când mușchii sunt sub stres), atunci aportul suplimentar este asigurat de metabolismul anaerob (suntem în condiții de lipsă de oxigen: dificultăți de respirație, oboseală în respirație etc.): este necesar să se accelereze acest metabolism prin conversia lactatului (care se obține din glicoliză) în glucoză prin gluconeogeneză.
Metabolismul aerob se dezvoltă în mitocondrii.
Prima enzimă întâlnită în metabolismul aerob este piruvat dehidrogenază; Este mai exact să spunem că piruvatul dehidrogenază este mai degrabă un complex enzimatic decât o enzimă, deoarece este un agregat de 48-60 unități proteice cu trei situsuri catalitice care acționează succesiv.
Piruvatul dehidrogenază catalizează următoarea reacție (redox):
Piruvat + NAD + + CoA-SH → Acetil CoA + NADH + H + + CO2
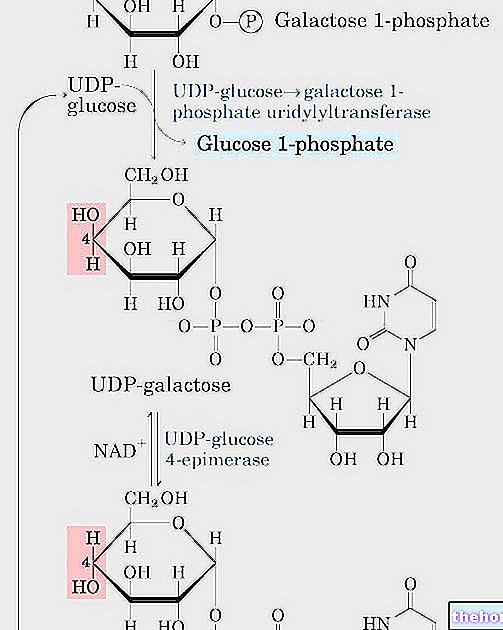
CoA-SH este coenzima A: este un derivat al acidului pantotenic; acetil coenzima A este un tioester. Acesta este un proces redox, deoarece primul carbon al piruvatului trece de la oxidarea numărul trei la oxidarea numărul patru (se oxidează) și al doilea carbon al piruvatului trece de la oxidarea numărul doi la oxidarea numărul trei (a oxidat). Apoi piruvatul este oxidat (pierde doi electroni în total) și NAD este redus.
După cum sa menționat, piruvatul dehidrogenază are trei tipuri de activitate enzimatică, fiecare susținută de propriul său cofactor catalitic:
- tiamina pirofosfat (este un derivat al vitaminei B1); este activ sub formă deprotonată: se formează un carbanion.
- lipoamida (este un derivat al acidului lipoic); conține o punte disulfură foarte reactivă.
- flavin adenin dinucleotid (este un derivat al vitaminei B2); este un nucleotid cu proprietăți redox: centrul său redox este alcătuit din flavină.
În celulele eucariote, metabolismul aerob apare în organele specializate ale celulei care sunt mitocondriile; la bacterii metabolismul glucozei și al altor specii are loc în celulă, dar nu există organite specializate.
Când piruvatul intră într-o mitocondrie, acesta este supus „acțiunii piruvat carboxilazei dacă este nevoie de efectuarea gluconeogenezei (pentru reconstituirea materialului de pornire) sau poate fi supus piruvat dehidrogenazei dacă este necesar pentru a produce energie: „acetil coenzima A care se formează prin metabolism aerob stimulează acțiunea piruvat carboxilazei, prin urmare, promovează gluoconeogeneza și reduce acțiunea piruvat dehidrogenazei.
Să vedem acum cum funcționează piruvatul dehidrogenază; în primul rând, există o decarboxilare a piruvatului prin acțiunea tiaminei pirofosfat.
Un mediu acid poate inhiba metabolismul aerob, deoarece forma anionică a tiaminei pirofosfat este activă, care ar fi protonată la pH acid și decarboxilarea nu ar avea loc.
O decarboxilare este o reacție dificilă, deoarece o legătură carbon-carbon trebuie ruptă; în acest caz, reacția este favorizată termodinamic de faptul că intermediarul de reacție (hidroxietil-tiamina pirofosfat) dă rezonanță (electronii p ai moleculei sunt delocalizați): hidroxietil-tiamina pirofosfatul există în trei forme posibile (de rezonanță) și acest lucru îl face destul de stabil. Mai mult, pirofosfatul de hidroxietil-tiamină sub formă anionică supraviețuiește suficient de mult timp pentru a putea interacționa cu puntea disulfură a lipoamidei (al doilea cofactor catalitic al piruvat dehidrogenazei); puntea disulfură este un braț oscilant (este situat la capătul unui lanț lung flexibil) și se poate deplasa de la un sit catalitic la altul din complexul enzimatic.
Apoi lipoamida, prin puntea disulfură, leagă pirofosfatul de hidroxietil-tiamină: se obține acetil lipoamida.Cel descris tocmai este prima etapă a unei reacții de transacetilare catalizată de prima enzimă a complexului piruvat dehidrogenază; în această fază s-a rupt o legătură între gruparea hidroxil și tiamina pirofosfat care a revenit la forma inițială: a avut loc o reacție redox în care puntea disulfură a acționat ca oxidant (cei doi atomi de sulf reduși) în comparațiile din grupul hidroxil care oxidat la acetil.
După această fază, brațul oscilant al lipoamidei se mișcă și se apropie de a doua enzimă a piruvat dehidrogenazei care desfășoară adevărata activitate transacetilază purtând grupa acetil cu ea: are loc a doua fază a reacției de transacetilare catalizată de a doua enzimă; în acest mod am obținut acetil coenzima A. Acum este necesar să restabilim lipoamida care este în formă redusă: intervine a treia enzimă a piruvat dehidrogenazei care redoxoxează lipoamida și își transferă electrozii către FAD care este redus la FADH2. FAD / FADH2 poate funcționa ca o pereche redox în două etape monoelectronice distincte sau într-o singură etapă bielectronică.
FADH2 își dă imediat electronii NAD + obținând FAD și NADH + H +.
Acetil coenzima A, obținută așa cum este descrisă, este produsul de pornire pentru ciclul Krebs (sau ciclul acizilor tricarboxilici).